Habitat/Ökologie (Meinunger & Schröder 2007) Amblystegium varium ist eine gut abgegrenzte Art, mit deren Bestimmung sich viele Bryologen schwer tun. Man muss die Art an richtig bestimmtem Vergleichsmaterial kennen lernen. Bestimmungssicherheit gewinnt man erst dann, wenn man die nächstverwandten Artgruppen, insbesondere A. serpens, A. juratzkanum, A. tenax, A. humile und A. trichopodium in allen ihren Erscheinungsformen gut kennt. Erschwerend kommt hinzu, dass alle diese Arten oft in Mischrasen durcheinander wachsen und oft einzeln voneinander getrennt werden müssen. Wie bei allen schwierigen Arten darf man nicht Einzelmerkmale isoliert für sich betrachten, es muss die Kombination wichtiger Merkmale zusammenstimmen und korreliert sein. Beispielsweise ist das oft bemühte Merkmal der geknieten Blattrippe zwar richtig, genügt aber für sich genommen nicht zur sicheren Bestimmung, da auch A. juratzkanum und A. trichopodium gekniete Rippen haben. Auch die neuere gängige Bestimmungsliteratur, etwa Smith (1980 und spätere Auflagen), Frahm & Frey (1983 und spätere Auflagen) und Nebel & Philippi (2001) ist nur mit Einschränkungen verwendbar. Völlig abwegig ist die Darstellung bei Touw & Rubers (1989), die man nicht benutzen sollte, hier werden auf Seite 407 unter Nr. 106.3 A. varium, A. radicale und A. humile, drei wirklich gut unterscheidbare Arten, zusammengefasst, was absolut inakzeptabel ist. Nach unseren Erfahrungen finden sich die besten Darstellungen bei Loeske (1903) und Nyholm (1979), die man vorrangig benutzen sollte. Wie bei allen schwierigen pleurokarpen Moosen sind zur sicheren Bestimmung gut entwickelte Stammblätter erforderlich. Astblätter oder kümmerlich entwickelte Pflanzen sind ungeeignet und sollten besonders am Anfang unbeachtet bleiben. Amblystegium varium ist A. serpens s. str. sehr ähnlich und bildet grüne, mehr oder weniger dichte Rasen mit anliegenden oder an der Spitze etwas abstehenden Blättern. Loeske (1903) beschreibt die Art sehr zutreffend: „Etwas kräftiger als A. serpens, von dem es durch die am Grund herzeiförmigen Blätter, die längeren Rippen und das meist sehr regelmäßig kurz parenchymatischsechsseitige Zellnetz mit derberen Wänden zu unterscheiden ist“. Bei A. serpens s. str. ist die Rippe immer gerade und endet in der Blattmitte oder kurz darüber. Amblystegium varium hat eine kräftige Rippe, die oben knieförmig verbogen ist und bis in die Blattspitze geht. Auch im oberen Blattteil ist die Rippe immer gut von den kurzen Laminazellen abgegrenzt. Bei A. juratzkanum und bei den trichopodium-Formen von A. humile (siehe dort) gehen die Rippen ebenfalls in die Blattspitze und sind verbogen, sie verlieren sich dort jedoch diffus in den meist länger gestreckten Laminazellen. Die Blätter der beiden letztgenannten Arten sind länger und schmaler und am Grunde nicht herzförmig verbreitert, besonders die Astblätter oft etwas gezähnelt, während die Blätter bei A. varium immer ganzrandig sind. Kümmerformen von Cratoneuron filicinum werden oft mit Amblystegium varium verwechselt, hier muss man sorgfältig an den Stammblättern nach den für C. filicinum typischen dünnwandigen, hyalinen, scharf abgesetzten Blattflügelzellen suchen. Amblystegium tenax unterscheidet sich durch kräftigere, nicht knieförmig verbogene und an der Blattspitze meist etwas austretende Rippe sowie andere Sporen, vergl. Nyholm (1979). Auf feuchten bis nassen, meist kalkhaltigen oder neutralen Unterlagen. Regelmäßig auf Steinen, Rinde und Wurzeln im Überschwemmungsbereich großer Flüsse, hier als Begleiter im Tortulo-Leskeetum polycarpae mit Leskea polycarpa, Leptodictyum riparium, Amblystegium fluviatile, A. serpens, Cratoneuron filicinum, Brachythecium rivulare, gesicherte soziologische Aufnahmen bei Ahrens (1992). Außerhalb der Flusstäler an kalkhaltigen Quellstellen, an Teich- und Seeufern und an feuchten Felsen in Neckerion-Gesellschaften. Wegen großer Bestimmungsunsicherheiten in der Vergangenheit muss die Soziologie erst noch neu erarbeitet werden.
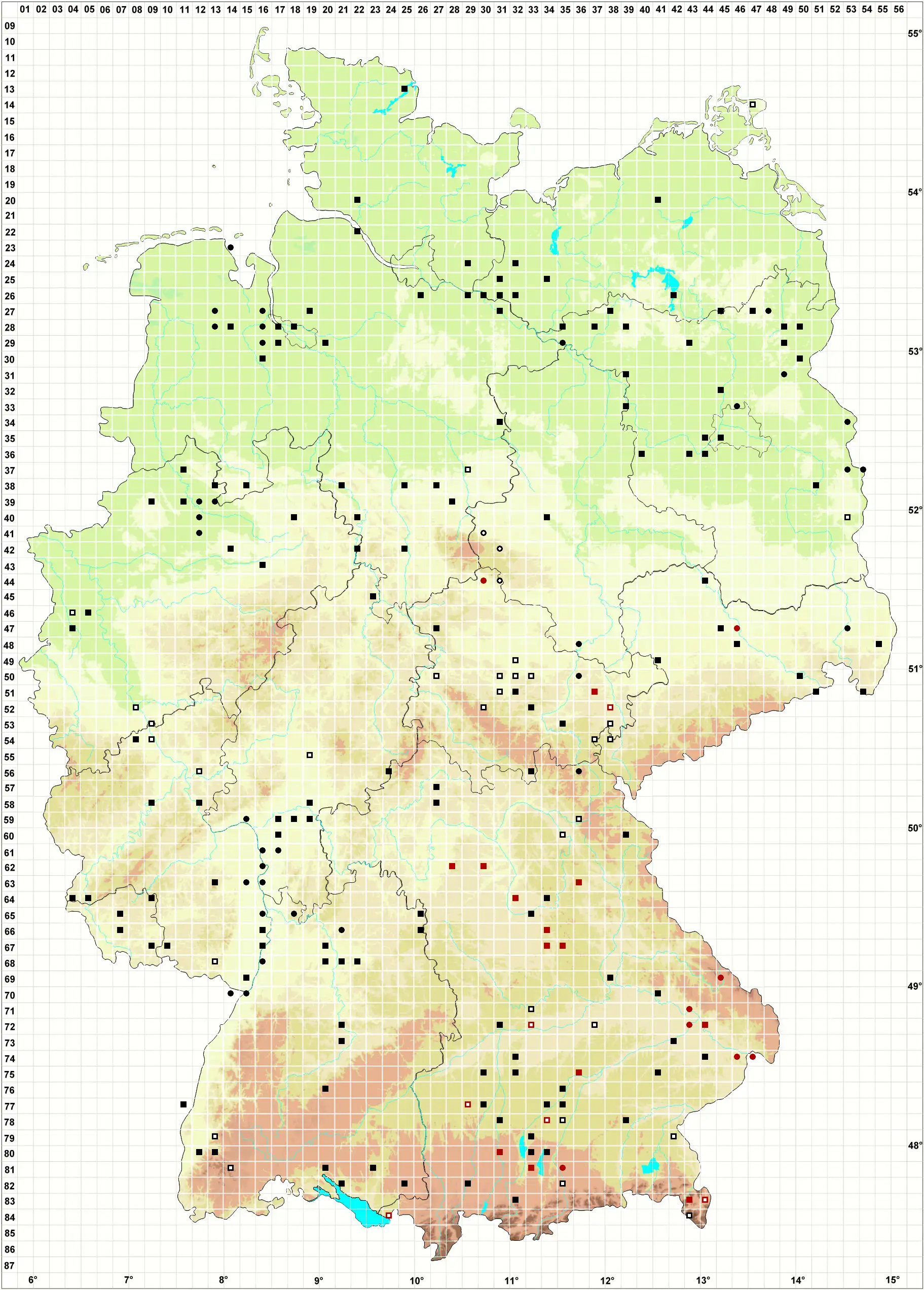
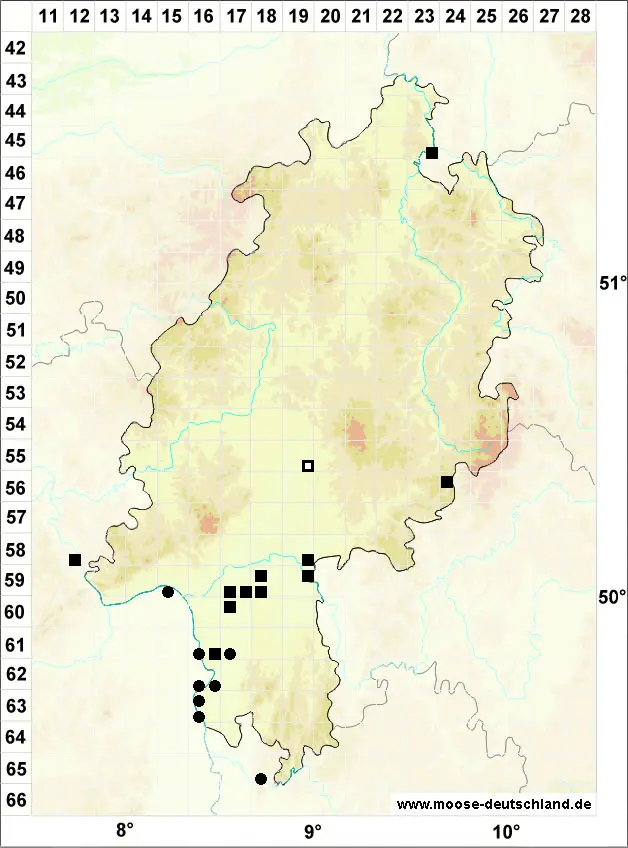
Verbreitung (Meinunger & Schröder 2007) Zerstreut, aber im ganzen Gebiet. Hauptverbreitung im Flach- und Hügelland und in den großen Flusstälern, geht aber vereinzelt auch in höhere Lagen. In die Karte wurde fast nur von uns geprüftes Material übernommen. Neben eigenen Funden und Belegen zahlreicher Mitarbeiter haben wir das Material in JE und REG durchgesehen. Literaturangaben blieben wegen großer Unsicherheiten weitgehend unberücksichtigt. SH: 1325/3 Schleiufer SW Karschau, mit Drepanocladus aduncus, 26.03.1999, WS; 2222/1 Steine am Elbufer bei Glückstadt, 15.07.1994, WS; 2429/3 Elmenhorst, am Weg nach Sahms, c. spor., 13.06.1947, G. Mechmershausen (!). MV: 2642/2 Pappelwald N Rechlin, 01.10.1995, WS; Linke et al. (1998a); E. Walsemann (!); Belege JE (!). NE: Koperski (1999 und weitere Belege !); Buse (1998); T. Homm (!). BB: 2837/2 quelliger Erlenwald NW Seddin, an Stein, 02.10.1998, LM; J. Klawitter (!); D. Benkert (!); S. Rätzel (!); S. Runge (!); F. Fuchs (!); Belege JE (!). NB: 4225/3 Ducksteinquelle NO von Lutterbeck, auf Kalksteinplättchen im Buchenwald, 1987, E. Volger (!); R. Düll (!); M. Koperski (!); G. Waesch (!); F. Jurkutat (!). ST: Loeske (1903); Marstaller (2001b); M. Koperski (!); Belege in JE (!). NW: Solga (1998); R. Düll (!); C. Schmidt (!); Belege in JE (!). HE: Manzke (1993 und weitere Belege !); T. Wolf (!); M. Preussing (!); Belege in JE (!). TH: Loeske (1903); 4727/3 Schlossberg NW Lengenfeld, Kalkblock im mesophilen Wald, 01.11.2000, M. Preussing (!); K. Strumpf (!); zahlreiche Belege JE (!). SN: M. Reimann (!); F. Müller (2004, !); Belege in JE (!). RP, SL: 6915/2 alte Tongrube S Jockgrim, 15.05.1995, LM; Lauer (2005); Oesau (2001); R. Düll (!); R. Mues und Mitarbeiter (!); S. Caspari (!). BW: In der Rheinebene und am Neckar anscheinend ziemlich verbreitet. Die Art geht aber bis in den Hochschwarzwald: 6526/1 Wald S von Klingen, Kalkstein, 14.04.1995, LM, 7620/3 Jurafelsen unter Burg Eineck, 15.08.1999, WS; 8013/3 Im Langenbach NW Schauinsland, 800 m, 04.09.1998, WS; 8225/4 Neuweiher NW Siggen, Uferzone, 08.09.1999, WS; M. Ahrens (1992; 1995 und weitere Belege !); U. Schwarz (!); U. Koch (!); M. Reimann (!); mehrere Belege JE, u. a. 8114/1 Baumstumpf am Ufer des Feldsees, 1120 m, Juli 1898, T. Herzog (!). BY: Nordbayern: 6228/3 Schellenberggraben S Castell, mit Brachythecium populeum, 24.02.1992 (!); 6230/3 Erlenbruch am Winterbach N Frimmersdorf, 11.05.2000, WS; 6626/4 Kalkstein im Wald S Bettenfeld, 16.04.1995, LM; 6735/2 Deusmauer Moor, Erlenbruch, 26.04.1997 (!!); 6938/3 Steinbruch N Kleinprüfening, 5/1993, J. Klotz & O. Dürhammer (!); M. Reimann (!); 6434/2 zwischen Hörnbach und Griesmühle bei Rupprechtstegen, 25.04.1997, B. Kaiser (!); K. Offner (!); Belege aus 6035/3+4 im Herbar Kaulfuss, UBT (!); Marstaller (2002); Belege in JE (!). Südbayern und Alpen: 7444/2 in Gräben N von Aidenbach und O Bergham, 10.03.1998, WS; 7532/4 Teichgelände W Blumenthal, 23.04.1997, WS; 7635/3 Amper S von Fahrenzhausen, 01.04.1998, WS; 7730/3 Guggenberg, 15.04.1997, WS; 7839/1 N von Permanöd, sumpfiger Waldbach, 30.03.1998, WS; 7933/3 S von Wörthsee, 21.05.1998, WS; 8031/2 Moor NW Oberhausen, 18.08.1996, WS; 8133/2 S Bernried, auf Ufersteinen am Starnberger See, 16.08.1996, WS; 8229/4 Sumpfgebiet W Sulzschneid, 18.05.2005, K. Offner (!); 8332/1 Bayersoier See, Baumstumpf am Wasser, 13.10.1994, R. Lotto (!); 8343/4 Berchtesgaden, Schapbach, 800 m, 30.06.1988, U. Beyerlein (!); 8235/2 Greilinger Moor bei Tölz, 4/1912, Hammerschmid (REG, !); 8443/2 Nordrand des Königsee, leg. Molendo 1859, det. LM (REG, !); Belege in JE (!).
Bestand und Gefährdung (Meinunger & Schröder 2007) Amblystegium varium findet sich meist in kleineren Rasen und als Begleiter zwischen anderen Moosen. Die Gefährdung der Art im Gesamtgebiet dürfte, wenn überhaupt, nur gering sein, doch sind gesicherte Aussagen wegen mangelhafter Datenlage derzeit nicht möglich. Umfangreiche Herbarauswertungen müssen hier Klarheit bringen.

 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]