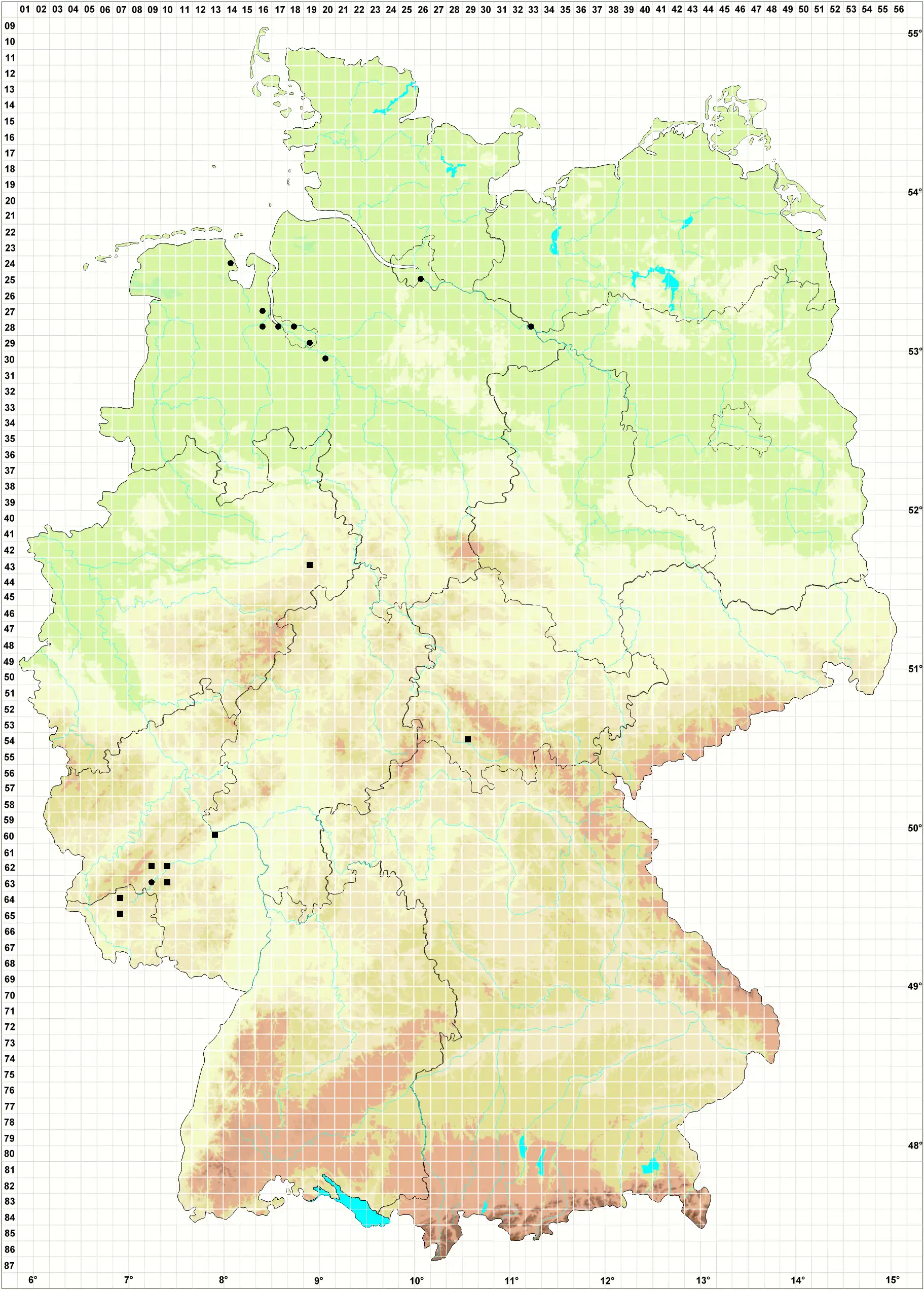
Habitat/Ökologie (Meinunger & Schröder 2007) Die Art wurde erst in jüngster Zeit für das Gebiet nachgewiesen. Nachträglich stellte sich heraus, dass sie bereits seit etwa 1970 von mehreren Bryologen hier gesammelt, aber nicht erkannt worden war. Scleropodium cespitans ist offenbar vielgestaltiger, als dies in der älteren Literatur dargestellt wurde. Koperski (1996c) und Smith (2004) weisen auf große Ähnlichkeiten mit Rhynchostegium murale und Cirriphyllum crassinervium hin. Verwechslungsgefahr besteht weiter mit sterilen Formen von Rhynchostegium riparioides, die oft nur durch unterschiedliche Blattflügelzellen der Stammblätter sicher zu unterscheiden sind. Mc. Adam & Smith (1981) veröffentlichten Brachythecium appleyardiae als neue, angeblich im südwestlichen England endemische Art, die bei Smith (2004) erneut verschlüsselt und ausführlich beschrieben wird. Der Status dieser Pflanzen blieb lange zweifelhaft und umstritten. Um 1970 sammelte der Erstautor in Thüringen Proben, die er als Scleropodium cespitans bestimmte. Das Material wurde F. Koppe und R. Schumacker vorgelegt, die diese Bestimmung nicht akzeptierten und es als zu Rhynchostegium murale gehörig erklärten, was uns wenig überzeugte. Inzwischen kamen von anderen Bryologen einige weitere ähnliche Proben hinzu. Weitere Bestimmungsversuche führten zu der Vermutung, dass dieses Material zu B. appleyardiae gehören könnte, was schließlich auch von Smith bestätigt wurde, vergl. Schmidt & Heinrichs (1999). Die Angabe „Germany“ in Smith (2004) bezieht sich auf diese Fundstellen. In einer neuen Arbeit stellen Blockeel et al. (2005) Brachythecium appleyardiae in den Formenkreis von Scleropodium cespitans. Typisches B. appleyardiae ist durch schmal lanzettliche, kaum hohle, feucht aufrecht abstehende Astblätter charakterisiert, die Äste machen feucht einen etwas stachligen Eindruck. Extremformen von Scleropodium cespitans haben kürzere, deutlich hohle Blätter, die Äste erscheinen dadurch kätzchenförmig und sind Rhynchostegium murale ähnlich. Die meisten Proben nehmen eine Mittelstellung zwischen diesen beiden ein. Nach Durchsicht unseres Materials können wir die Ergebnisse von Blockeel et al. (2005) bestätigen. Wir führen diese beiden Arten (bzw. Formen) nicht mehr getrennt und fassen sie unter der Bezeichnung Scleropodium cespitans zusammen. Grüne, an schattigen Stellen blass- bis gelblichgrüne, oft ausgedehnte Rasen auf feuchten bis nassen kalkhaltigen Gesteinen, von da aus auf Erde und Baumwurzeln übergehend. Hauptverbreitung an meist nordexponierten Uferfelsen von Flüssen oberhalb der Mittelwasserlinie in Bereichen, die nur bei Hochwasser kurze Zeit überschwemmt werden. Seltener an Sickerwasserstellen von Felsen. Im Tiefland vorwiegend auf Gestein von Uferbefestigungen großer Flüsse und Kanäle, von da aus auf Weidengebüsche und Erde übergehend, selten auch am Grunde von Bäumen an trockeneren Stellen. Typische Begleiter sind: Rhynchostegium murale, R. riparioides, Brachythecium rivulare, B. rutabulum, Leptodictyum riparium, Homalia trichomanoides, Leskea polycarpa, Tortula latifolia, Cirriphyllum crassinervium, Didymodon sinuosus, D. insulanus, Eurhynchium praelongum und E. hians agg. Im Nahebergland an sickerfeuchten Felsen mit Homalothecium sericeum, Neckera besseri, Metaneckera menziesii. Soziologisch-ökologische Hinweise finden sich bei: Koperski in Ludwig et al. (1996); Sesterhenn & Caspari (1998); Caspari (2004).
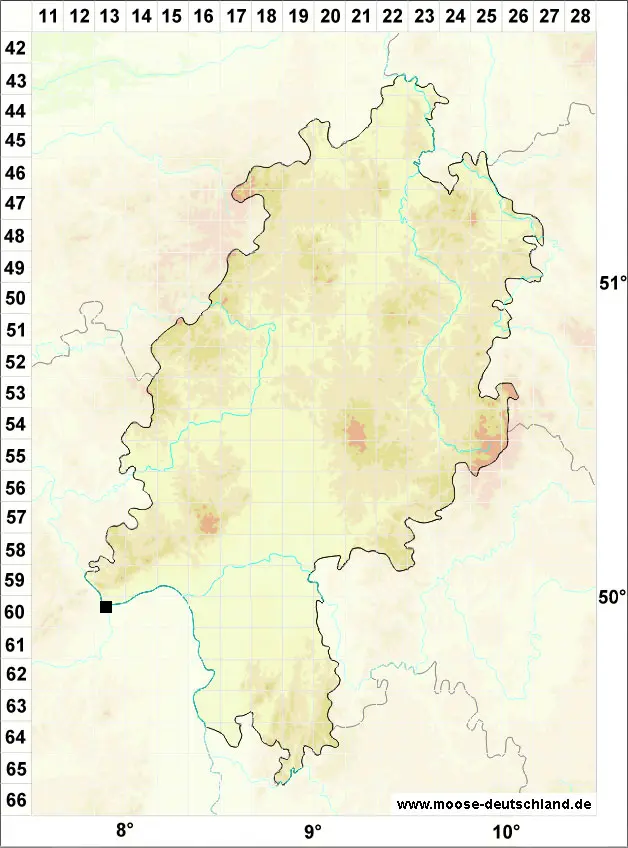
Verbreitung (Meinunger & Schröder 2007) Die ozeanisch-submediterrane Art befindet sich im Gebiet an ihrer östlichen Arealgrenze. Im nordwestdeutschen Flachland im Weser- und Elbebereich erst in neuerer Zeit eingewandert und anscheinend in weiterer Ausbreitung. Die Vorkommen an naturnahen Standorten im mittel- und westdeutschen Hügelland sind offenbar alteinheimisch und wurden früher nur übersehen. SH: 2526/1 NSG Heuckenlock bei Stillhorn, Frahm (2001a). – Zu streichen oder zweifelhaft: Zwei Angaben um Hamburg E. Walsemann in Koperski (1996c), ein uns vorliegender Beleg von Lankau ist ein steriles, nicht näher bestimmbares Brachythecium: LM (!). NE: Hauptsächlich an Uferbefestigungen im Weser- und Elbegebiet: Homm et al. (1995); Koperski (1996c; 1999); Koperski in Ludwig et al. (1996); Homm (1999); 2833/1 Schlosspark Wehningen, Stammfuß einer alten Eiche, 18.07.1995, M. Koperski (!), entspricht typischem Brachythecium appleyardiae. NW: 4319/1 Kreidekalkfelsen an der Sauer bei Grundsteinheim, 22.11.1995, C. Schmidt (!), vergl. Schmidt & Heinrichs (1999). TH: 5429/3 nordexponierte Muschelkalkfelsen an der Werra O von Vachdorf und an der Wasserleite zwischen Henfstädt und Leutersdorf, LM. Die Art wurde dort erstmalig um 1970 gesammelt, jedoch wegen Bestimmungsunsicherheiten bei Meinunger (1992) nicht aufgenommen. Bis heute reichlich. RP, SL: Mehrfach im Flussgebiet der Nahe und der Prims: Sesterhenn & Caspari (1998); Caspari et al. (2000); Caspari (2004); 6013/1 Bingen, östliche Seite der Nahemündung, auf Salix fragilis, 22.06.1998, leg. A. Oesau, det. S. Caspari (!); 6407/332 bei Büschfeld, am Prallhang zur Prims gegenüber dem Saargummiwerk, 26.11.2000, S. Caspari (!); 6407/323 NSG Bardenbacher Fels, 18.12.2000, S. Caspari (!); 6507/112 Prallhang zur Prims bei Büschfeld, 26.11.2000, S. Caspari (!)
Bestand und Gefährdung (Meinunger & Schröder 2007) Die Art kommt öfter in großen Beständen vor. In Norddeutschland hat sie sich in neuerer Zeit weiter ausgebreitet: Koperski (2006a), hier ist sie derzeit ungefährdet. Die sicher alteinheimischen Vorkommen an felsigen Steilhängen weiter südlich sind ebenfalls gut besetzt und nicht unmittelbar bedroht. Da es sich dabei immer um bryologisch reiche Sonderstandorte handelt, sollten sie alle, soweit noch nicht geschehen, als Schutzgebiete ausgewiesen werden. RL R.


 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]