Habitat/Ökologie (Meinunger & Schröder 2007) Gelblichgrüne, unregelmäßig gefiederte Pflanzen in lockeren, verworrenen Rasen oder zwischen anderen Moosen umherkriechend. Bei guter Entwicklung stehen die Blätter, besonders an den Sprossspitzen, sternförmigsparrig ab und geben der Art im Gelände ein charakteristisches Aussehen. Im außeralpinen Deutschland wächst (wuchs) die Art, wie Caspari (2004) sehr zutreffend schreibt, an Auflichtungsstellen des Galio-Carpinetum auf teilbeschattetem, grusigem Waldboden, insgesamt meist an trocken-warmen Standorten. Unterlagen sind schwach basische Silikatgesteine wie Porphyr, Basalt, Schiefer, Granit oder Gneis, aber auch Gips oder entkalkter Löß. Als „säureliebende Pflanze schattiger Standorte“, wie bei Nebel & Philippi (2001) angegeben, kann man die Art hier nicht bezeichnen. Die Wuchsstellen sind als neutral bis schwach sauer zu bezeichnen, die Art wächst (wuchs) gerne an steilen, felsigen Weganschnitten, die mit lockeren Schlehengebüschen bestanden waren zwischen lockerem Grasbewuchs aus Brachypodium pinnatum und Poa nemoralis. Wichtige Begleitmoose sind, wie auch Caspari (2004) angibt, Brachythecium velutinum und Hypnum cupressiforme filiforme. Sie scheint sehr konkurrenzschwach und sehr eutrophierungsempfindlich zu sein, beim Auftreten nitrophiler Arten verschwindet sie sofort. Soziologische Aufnahmen aus dem Gebiet liegen nicht vor. In den Alpen wächst die Art in Zwergstrauchheiden unter Krummholz und in alpinen Rasen, Grims (1999). Auch aus diesen Gebieten liegen nähere Angaben zu Ökologie und Soziologie nicht vor.
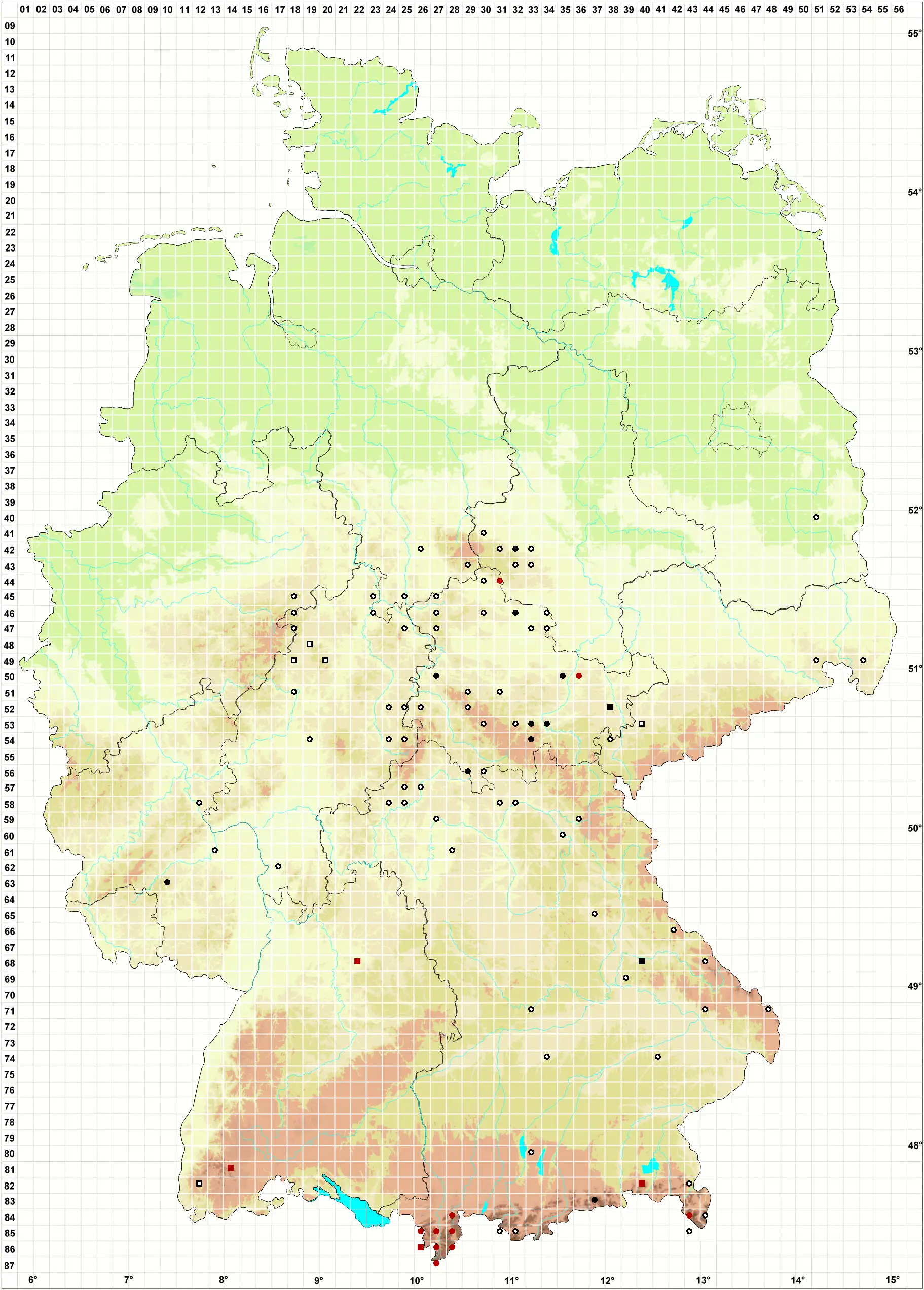
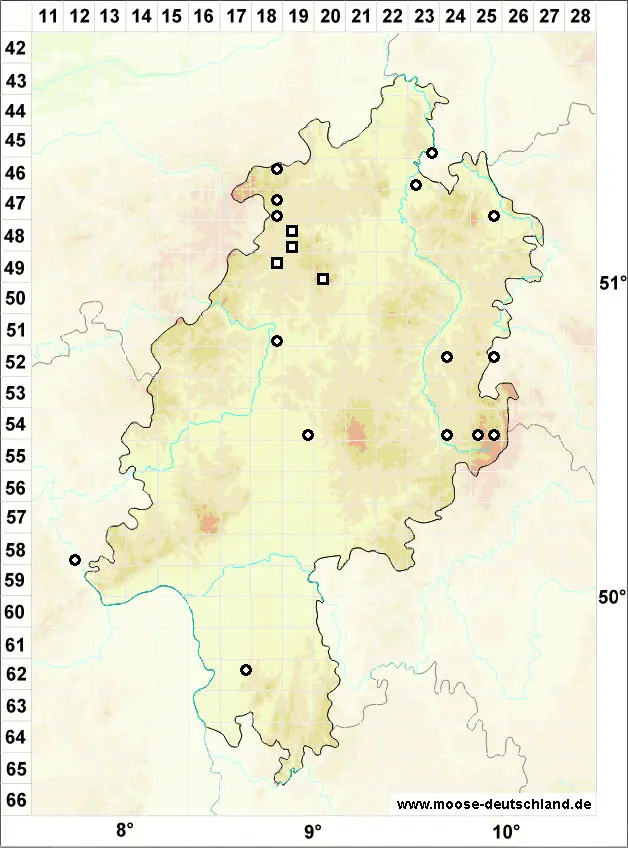
Verbreitung (Meinunger & Schröder 2007) Die Art wurde früher öfter falsch bestimmt, ihre Häufigkeit wurde auch früher vermutlich stark überschätzt, ungeprüfte Literaturangaben sind mit Vorsicht zu bewerten. In den Alpen wurde sie um die und oberhalb der Baumgrenze gefunden. In den übrigen Gegenden konzentrieren sich jedoch die Nachweise auf die submontane und colline Stufe in hainbuchenreichen, klimatisch eher subkontinental getönten Gebieten in der Mitte und im Osten: BB: 4051/2 bei Lieberose, Busch in Milde (1869), seitdem verschollen: Klawitter et al. (2002). NB: Quelle (1902), seitdem verschollen. ST: Mehrfach im Unterharz, Loeske (1903); Zschacke (1905); Nörr (1969). Keine Nachweise aus jüngerer Zeit. NW: F. Koppe (1977), verschollen, Schmidt & Heinrichs (1999). HE: Grimme (1936); Röll (1926); F. Koppe (1977). Im Herbar J. Futschig (FR !) befinden sich folgende Belege: 4819/1 Fürstenberg; Schmittlotheim-Kirchlotheim; 4819/3 Altenlotheim; 4918/2 N Röddenau; 4920/3 Dodenhausen. TH: Röll (1915); Reimers (1940; 1955); Fröhlich (1969); Meinunger (1992). Nachweise aus jüngster Zeit: F. Müller (1993a); 5036/1 NSG Gleistalhänge, Lößlehm über der Flachsleite und 4632/1 Westabfall der „Kammern“ SW vom Rathsfeld, 11/2004, R. Marstaller; 5238/1 Aumatal bei Weida und „Die Burg“, 10.04.1979, J. Rettig (!). SN: Milde (1869); Riehmer (1926); Reimann (1997); F. Müller (2004). Verschollen. RP: Caspari et al. (2000); zwei aktuelle Funde: 6310, Caspari (2004); Lauer (2005). – Zu streichen: Angaben Düll (1980; 1995). BW: Sicher belegt nur von zwei Stellen im Südschwarzwald, alle übrigen Literaturangaben sind zu streichen oder zweifelhaft: Nebel & Philippi (2001). BY: Familler (1913); Paul & Poelt (1950); Paul & v. Schoenau (1914; 1928; 1933); K. Koppe (1968); R. Lübenau (!). Nachweise aus jüngster Zeit: 6840/2 kleine Granitfelsen an der Straße N Trasching, mit Brachythecium velutinum und Hypnum cupressiforme, 09.08.1997, LM; 8627/3 Fellhorn, 05.09.1996 (!!); 8727/2 Kratzer, M. Reimann (!); 8526/4 und 8527/3, M. Preussing (in litt.). 8628/1 Osthang des Salober, 05.09.2004, M. Reimann (2005); 8337/4 Rotwand, Wegböschung oberhalb Rotwandhaus, 14.09.2004, Dürhammer & Reimann. – Zu streichen: F. & K. Koppe (1931), ist Heterocladium heteropterum, rev. M. Sauer (STU).
Bestand und Gefährdung (Meinunger & Schröder 2007) Von dieser schon früher im Gebiet nicht häufigen Art sind gegenwärtig kaum zehn existente Vorkommen bekannt, wobei die Bestände immer klein bis winzig sind. Nebel & Philippi (2001) vermuten als Ursache für diesen enormen Rückgang hauptsächlich Veränderung des Standortes durch Eutrophierung. Caspari (2004) kommt für die letzten Vorkommen in RP in 6310 Steinalptal zu ähnlichen Vermutungen: Diese liegen im Zentrum eines Truppenübungsplatzes, wo im Umkreis von mindestens 7 km keinen intensive Landwirtschaft stattfindet und die Verkehrsbelastung gering ist. Heterocladium dimorphum steht im Gebiet vor dem Aussterben: RL 1. Nur in den Hochlagen der Alpen erscheint die Situation etwas günstiger, hier ist derzeit eine Einstufung als RL R vertretbar.


 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]