Habitat/Ökologie (Meinunger & Schröder 2007) Eine atlantische Art, die erst in neuester Zeit in Deutschland nachgewiesen werden konnte. Die europäischen Areale von Scapania gracilis und Douinia ovata sind sehr ähnlich, siehe K. Müller (1951–1958). Beide Arten wurden bei uns im gleichen Gebiet gefunden. Auf Silikatfelsen, Holz und zwischen anderen Moosen. Cephaloziella Die Gattung Cephaloziella gehört zu den schwierigsten unter allen Moosen. Bis in die Gegenwart sind alle Lokalbearbeitungen durch Unsicherheit und Hilflosigkeit gekennzeichnet. K. Müller (1951–1958: 1018) schreibt dazu in seinem zusammenfassenden Werk: „Nur wer sich selbst schon eingehender mit diesen kleinsten Lebermoosen befaßt hat, kann ermessen, welche Unsumme von mikroskopischer Arbeit nötig war, um zu einem klaren Bild zu kommen. Es scheint mir ganz zwecklos, eine Pflanzenfamilie, die an sich schon von nur ganz wenigen Hepatikologen einigermaßen beherrscht wird, in eine Unmenge von Arten aufzusplittern, die den übrigen Arten dieses Handbuchs gar nicht gleichwertig sind und auch von kaum jemand auseinandergehalten werden können“. Von dieser Aussage ist nur die erste Hälfte richtig, die zweite nicht. Über diese unbefriedigende Situation haben in der Folgezeit vor allem die Arbeiten von Schuster (1980) und Damsholt (2002) wesentlich hinausgeführt. Wir haben im Laufe der Jahre mehrere tausend Proben untersucht. Neben eigenen Aufsammlungen und Proben zahlreicher Mitarbeiter wurden die Herbarien in DR, JE, REG, die Sammlung K. Koppe in HAL, die Sammlung F. Koppe in STU sowie das Herbar J. Futschig in FR ausgewertet. Besonderer Dank gilt Uwe Schwarz, der sein umfangreiches Herbar zur Verfügung stellte und schwer zugängliche Literatur beschaffte. Wir sind zu dem Ergebnis gekommen, dass sich im Gebiet die nachfolgend genannten Arten sehr wohl unterscheiden lassen und auch eine eigenständige Ökologie zeigen, die künftig genauer untersucht werden muss. Mehr als in anderen Gattungen sind die Arten durch eine Summe von Merkmalen charakterisiert, die in ihrer Gesamtheit, oder, wie Schuster zutreffend schreibt, „in concert“ (ohne Disharmonien) zusammenstimmen müssen. Die auf den ersten Blick auffälligsten Merkmale sind oft nicht die entscheidenden, die Bestimmung hat gewissermaßen ganzheitlich „von innen heraus“ zu erfolgen. Man muss jetzt nicht mehr die Aussagen der Spezialisten ungeprüft und gläubig hinnehmen. Jeder gewissenhafte Bryologe, der systematisch und mit der nötigen Ausdauer die Probleme angeht, kann sich künftig die Gattung selbst erarbeiten. Der verständliche Wunsch aller Floristen, vor allem den Seltenheiten nachzuspüren, hat hier am Anfang gänzlich zurückzustehen, man muss zuerst die häufigen Arten, insbesondere Cephaloziella divaricata und C. rubella, in allen ihren Erscheinungsformen wirklich kennen und beherrschen lernen. Für eine erfolgreiche Beschäftigung mit der Gattung ist außer den eingangs genannten Hauptwerken folgende Literatur unerlässlich: K. Müller (1905–1916); Douin (1928); Paton (1999); Schumacker & Váña (2000). Man sollte seine Proben mit allen diesen Schlüsseln versuchen zu bestimmen, die oft auf unterschiedlichen Wegen zum gesuchten Ergebnis führen. Sehr zu empfehlen ist es, sich zahlreiche Dauerpräparate anzufertigen, die rasche Vergleiche nach allen Richtungen ermöglichen und allmählich den Blick für das Wesentliche schärfen, ohne dieses Hilfsmittel ist besonders am Anfang oft kein Durchkommen. Man beginne mit gut entwickeltem, möglichst fertilem Material. Zu beachten ist, dass Mischproben häufig sind, besonders Cephaloziella divaricata und C. rubella wachsen oft gemeinsam und durcheinander, die Proben müssen daher zunächst nach dem Augenschein vorsortiert werden. Als grundlegend wichtiges Bestimmungsmerkmal ist vor allem die Geschlechtsverteilung festzustellen. Es gibt diözische, autözische und parözische Arten. Die einzige diözische Art im Gebiet ist Cephaloziella divaricata, sie ist gleichzeitig hier die häufigste. Meist tritt sie entweder ganz steril oder in rein männlichen oder rein weiblichen Rasen auf. Glücklicherweise ist sie durch die kleinen Zellen und stets vorhandenen Unterblätter gut charakterisiert und kann auch ohne Feststellung der Diözie sicher bestimmt werden. Autözische Arten geben sich bei guter Entwicklung durch das gleichzeitige Vorhandensein von Perianthien und männlichen Trieben zu erkennen. Dabei sind die letzteren, auch wenn die Antheridien bereits ausgefallen sind, durch die vergrößerten, oft auch anders gefärbten Hüllblätter als ährenförmige Gebilde, die endständig (terminal) oder durchwachsen (intercalar) sein können, auffällig. Am schwierigsten sind parözische Arten zu erkennen. Nur bei sehr gutem Material findet man unterhalb der Perianthien in den vergrößerten und am Grunde meist etwas ausgehöhlten männlichen Hüllblättern noch Antheridien. Meist entwickeln sich zunächst die Antheridien, die dann später, wenn das Perianth ausgebildet ist, bereits ausgefallen sind. Oft kommt das Perianth nicht zur Entwicklung, und es sind nur die männlichen Ähren zu erkennen, die eine andere Geschlechtsverteilung vortäuschen können. Wichtig für die Bestimmung sind gut entwickelte sterile Triebe ohne Gemmen. Man findet sie meist nur an Material, das unter gleichmäßigen humiden Bedingungen gewachsen ist. Kümmerformen bestehen oft aus einem Gemisch von Gametangienständen in unterschiedlichen Entwicklungsphasen und gemmentragenden sterilen Sprossen. Sie sind zur Bestimmung ebenso wenig geeignet wie nass gewachsene Formen mit meist sehr entfernt stehenden, aber oft untypisch entwickelten Blättern. Die Triebe sind auf das Vorhandensein von Unterblättern zu untersuchen. An gut entwickelten Blättern im Mittelteil dieser Triebe ist die Blattform zu beurteilen und sind an der Basis der Blattlappen die Zellbreiten zu messen, die wichtige Bestimmungsmerkmale abgeben. Die sonstigen Merkmale sind meist stark veränderlich, und man muss ihre Toleranzbereiche durch Untersuchung zahlreicher unter verschiedensten Bedingungen gewachsener Proben kennen und abschätzen lernen. Zellwandverdickungen und Papillosität sind in weiten Bereichen und oft an einer Pflanze variabel. Meist zeigen die oberirdischen und rascher Austrocknung ausgesetzten Pflanzenteile dickwandige Zellen, während diese an den geschützteren Teilen weiter unten zunehmend dünnwandig werden. Form und Zähnung der weiblichen Hüllblätter können ebenso in weiten Bereichen variieren wie die Ausbildung der Perianthmündungszellen, die stark vom jeweiligen Entwicklungsstand abhängig ist. Das gleiche gilt für alle sonstigen Merkmale wie etwa Farbe der Brutkörper oder Färbung der Pflanzen, die sämtlich als „Merkmale zweiten Grades“ einzustufen und entsprechend vorsichtig zu verwenden sind. Wir geben nachfolgend einen verkürzten Schlüssel für die Arten des Gebietes, der nicht jede Ausnahme berücksichtigen kann, einige Vorkenntnisse voraussetzt und nur im Zusammenhang mit der oben genannten Literatur erfolgreich benutzt werden kann: Schlüssel für die Arten der Gattung Cephaloziella 1 Gemmen zweizellig, kubisch, eckig oder mit warzigen Erhebungen C. integerrima – Gemmen zweizellig, elliptisch, glatt, oder Gemmen fehlend 2 2 Diözisch, oft steril. Unterblätter deutlich. Blattzellen klein, 8–12 µm. Häufigste Art der Gattung C. divaricata – Monözisch, meist fertil 3 3 Nur auf schwermetallhaltigen Unterlagen, meist an alten Erzbergwerken. Blätter meist deutlich bis stark gezähnt, oft auch der Blattrücken. Sehr seltene Arten 4 – Auf verschiedenen Unterlagen. Blätter ganzrandig oder am Rande manchmal mit einzelnen Zähnen. Weiter verbreitete Arten 6 4 Zellen groß, 15–20 µm breit. Blätter vereinzelt und spärlich gezähnt. Parözisch C. nicholsonii – Zellen kleiner. Blätter stark bis dornig gezähnt. Autözisch 5 5 Zellen klein, 7–10 µm, oft stark verdickt. Blätter, Unterblätter und weibliche Hüllblätter mehr oder weniger stark gezähnt. Gemmen in der Mitte eingeschnürt C. massalongoi – Zellen mittelgroß, 12–15 µm. Blätter, Blattrücken, Unterblätter und weibliche Hüllblätter stark bis dornig gezähnt, igelstachelig. Gemmen in der Mitte nicht eingeschnürt C. phyllacantha 6 Parözisch 7 – Autözisch 14 7 Sterile Sprosse mit deutlichen, oft großen und zweiteiligen Unterblättern 8 – Sterile Sprosse ohne Unterblätter, gegen die Spitze hin gelegentlich mit zwei- bis dreizellreihigen, stummelförmigen Auswüchsen 12 8 Zellen groß, 14–20 µm breit 9 – Zellen kleiner und schmaler 10 9 Blätter wenigstens teilweise entfernt gezähnt, Apikalzellen verlängert. Blätter dem Stängel mehr oder weniger anliegend. Nur auf Schwermetallböden C. nicholsonii – Blätter ganzrandig, schräg und sparrig vom Stängel abstehend, weibliche Hüllblätter teilweise sternförmig ausgebreitet C. stellulifera var. stellulifera 10 Zellen klein, 7–10 µm breit. Blattlappen an der Basis 5–12 Zellen breit. Unterblätter außerhalb der Gametangienstände unscheinbar, aber meist regelmäßig vorhanden C. elegans – Zellen mittelgroß, 10–15 µm breit. Blattlappen an der Basis schmaler, 4–9 Zellen breit 11 11 Unterblätter außerhalb der Gametangienstände klein und unscheinbar. Pflanzen hell- bis gelbgrün, an sonnigen Stellen auch bräunlich, nur die Perianthien und die männlichen Hüllblätter gelegentlich leicht rosarot angelaufen. Ähnlich C. varians, aber parözisch C. stellulifera var. limprichtii – Unterblätter überall sehr groß und deutlich, tief zweiteilig. Pflanzen reingrün, Sprossspitzen und teilweise die Blattspitzen tief purpurrot gefleckt. Pflanze im Aussehen wie C. rubella var. rubella, die jedoch keine Unterblätter besitzt C. arctogena 12 Zellen klein, 7–10 µm breit, meist rundlich. Blattlappen an der Basis 5–12 Zellen breit. Sterile Sprossteile meist mit unscheinbaren, aber deutlichen Unterblättern C. elegans – Zellen mittelgroß, 10–16 µm breit, im unteren Teil der Blattlappen regelmäßig rechteckig geformt. Blattlappen an der Basis nur 4–5 Zellen breit. Gut entwickelte sterile Sprosse ohne Unterblätter. C. rubella agg. 13 13 Weibliche Hüllblätter oft weit hinauf zu einem Ring verwachsen, der den Grund des Perianths urnenförmig umschließt C. rubella var. rubella – Weibliche Hüllblätter tiefer gespalten und fast frei, die Basis des Perianths nicht urnenförmig umfassend C. rubella var. sullivantii 14 Sterile Sprosse ohne Unterblätter 15 – Sterile Sprosse mit deutlichen Unterblättern, mindestens in der Nähe der Sprossspitzen 20 15 Zellen groß, 14–20 µm breit 16 – Zellen mittelgroß, 10–15 µm breit 18 16 Weibliche Hüllblätter mit abgerundeten, ganzrandigen, oft etwas welligen Lappen, Perianth birnförmig, frei C. integerrima – Weibliche Hüllblätter mit spitzen, mehr oder weniger gesägten Lappen 17 17 Pflanzen hell- bis blassgrün, an sonnigen Stellen ± bräunlich. Zellen immer dünnwandig bis mäßig verdickt. Blattlappen an der Basis 6–10 Zellen breit C. hampeana var. subtilis – Pflanzen wenigstens teilweise rot, besonders die Antheridienstände oft tief purpur- bis schwarzrot. Zellen dünn- oder dickwandig. Die weiblichen Hüllblätter oft grob gesägt mit teilweise hakenförmig zurückgekrümmten Zellen. Perianthmündungszellen lang ausgezogen. Blattlappen an der Basis 4–6 Zellen breit C. rubella var. pulchella 18 Wärmeliebende Kalkpflanze, im Gebiet bisher nicht nachgewiesen, erst dicht jenseits der Grenze in Holland nachgewiesen C. baumgartneri – Auf kalkfreien Böden und zwischen anderen Moosen. Im Gebiet zerstreute Arten 19 19 Pflanzen blassgrün, an offenen Stellen teilweise bräunlich. Zellen immer dünnwandig. Blattlappen an der Basis 6–10 Zellen breit. Perianthmündungszellen bei gut entwickeltem Material teilweise bis zur Hälfte der Zelllänge fingerförmig frei, die Perianthmündung dadurch gezähnelt erscheinend C. hampeana – Pflanzen mehr oder weniger rot überlaufen, besonders die Hüllblätter der Antheridien meist kräftig rotbraun. Zellen teilweise deutlich bis stark verdickt. Blattlappen an der Basis 4–6 Zellen breit. Perianthmündungszellen auch bei gut entwickeltem Material fast bis oben verwachsen, die Perianthmündung dadurch gekerbt erscheinend C. rubella var. bifida 20 Zellen groß, 14–20 µm breit 21 – Zellen mittelgroß bis klein, 7–14 µm breit 22 21 Blattlappen sehr schmal und spitz, an der Basis 2–3 Zellen breit. Blätter an der Basis oft mit einzelnen, großen, seitlichen Zähnen. Hüllblätter der Gametangienstände meist mit großen, teilweise zurückgekrümmten Zähnen. Nur in Hochmooren und auf Torf C. elachista – Blattlappen an der Basis 5–10 Zellen breit, vorne abgerundet oder stumpflich gespitzt. Blätter ganzrandig. Weibliche Hüllblätter nur gekerbt oder schwach gezähnelt. Meist große Teile der Pflanze sowie die Brutkörper weinrot gefärbt C. grimsulana 22 Blattlappen an der Basis meist 2–5 Zellen breit, meist lang und schmal, bisweilen mit einzelnen Zähnen. Zellen meist deutlich, teilweise stark verdickt, oft papillös. Ältere und sonnig gewachsene Teile der Pflanze oft auffällig kupferfarbig. Kalkmeidend, in Mooren, auf Torf und feuchtem, kalkfreien Sand C. spinigera – Blattlappen an der Basis 6–12 Zellen breit, breiter und kürzer als bei voriger Art, meist ganzrandig. Zellen (im Gebiet) meist dünnwandig bis mäßig verdickt. Pflanzen grün, gelblich, braun oder rot, nicht auffällig kupferfarbig. An basenreichen, mehr oder weniger kalkhaltigen Standorten 23 23 Weibliche Hüllblätter auffällig stark gezähnt (ähnlich C. elachista), an der Spitze oft 2–3 verlängerte Zellen übereinanderstehend, seitliche Zähne an der Basis teilweise mehrzellig. Perianthmündungszellen oft über die Hälfte frei, teilweise ganz freistehend, zur Spitze hin stumpf dreieckig verschmälert. Gut entwickelte sterile Triebe immer mit großen, auffälligen Unterblättern C. uncinata – Weibliche Hüllblätter ganzrandig, gekerbt oder schwach gezähnt. Perianthmündungszellen meist weit hinauf verwachsen, an der Spitze oval abgerundet, nicht auffällig dreieckig. Unterblätter oft schwächer entwickelt, oft nur in der Nähe der Sprossspitzen vorhanden (C. varians agg.) 24 24 Stängel an der Spitze teilweise mit einzelnen hyalinen Zähnen, Blätter z. T. gezähnelt. Zellen an der Blattbasis am Rücken teilweise mamillös oder konisch aufgewölbt C. varians var. scabra – Stängel und Blätter glatt, ohne solche Auswüchse 25 25 Blattlappen eiförmig, an der Spitze abgerundet. Antheridienstände drehrund beblättert, dadurch kolbenförmig C. varians var. arctica – Blattlappen zugespitzt, an der Spitze oft mit zwei übereinanderstehenden Zellen. Antheridienstände meist deutlich einseitswendig beblättert (C. alpina Douin) C. varians var. varians Im anschließenden speziellen Teil bleibt die bisherige Literatur wegen großer Unsicherheiten weitgehend unberücksichtigt. Die meisten Verbreitungskarten wurden nur nach von uns geprüften Belegen gezeichnet und sind daher entsprechend unvollständig. Bei den im Text genannten Einzelfunden sind die wenigen, von anderen Autoren übernommenen Angaben klar gekennzeichnet. Alle übrigen Angaben wurden von LM bestimmt, überprüft oder revidiert, was nicht nochmals besonders erwähnt wird. - Cephaloziella baumgartneri Schiffn. Mediterranatlantische Art an sehr warmen, kalkhaltigen Standorten. Im Gebiet bisher nicht bekannt, wurde jedoch neuerdings dicht jenseits der Grenze in Holland nachgewiesen: During et al. (1986). Die Art ist vielleicht auch bei uns im Verbreitungsgebiet von Leiocolea turbinata noch zu erwarten. Ausführliche Beschreibungen der Pflanze finden sich bei K. Müller (1951–1958); Gradstein & van Melick (1996) sowie Paton (1999).
Verbreitung (Meinunger & Schröder 2007) Bisher nur einmal gefunden: NW: 4617/3 Bruchhauser Steine, ein kleiner Rasen, mit Paraleucobryum longifolium, Andreaea rupestris, Diplophyllum albicans und Anastrophyllum minutum, leg. H. Müller um 1858, det. C. Schmidt, Beleg in WWU-Münster (!). Der Beleg trägt die Aufschrift „139. Bruchhauser Steine“ in der Handschrift von H. Müller. Darunter ist zu lesen: „Scapania Bartlingii, Saroscyphus Ehrharti“ in der Handschrift von Beckhaus, beide Namen wurden aber wieder gestrichen. Schließlich ist in der Handschrift von F. Koppe vermerkt: „Scapania compacta (Roth) Dum.“, und diese falsche Bestimmung wurde in F. Koppe (1977) veröffentlicht. Anlässlich einer erneuten Revision des Materials erkannte C. Schmidt die Pflanzen als zu S. gracilis gehörig, was wir bestätigen konnten. Angaben zu H. Müller-Lippstadt finden sich in Frahm & Eggers (2001).


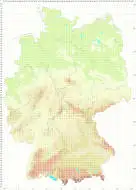 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]